
















Eléments fondamentaux et conséquences pratiques sur la gestion des maladies du gazon – R. Latin, Ph.D
Article original :
“Understanding Fungicide Resistance, R. Latin, 2017, Green Section Record. Vol. 55(13)1:7”
Traduit par R. GIRAUD avec la permission de l’USGA Green Section Record.

Cet article est téléchargeable au format *.pdf en cliquant sur le lien suivant.
Quiconque travaille dans l’entretien des pelouses devrait connaître le principe de « résistance aux fongicides ». Dans bien des cas, la résistance est un facteur de la mauvaise efficacité des fongicides. Ce phénomène a lieu lorsqu’une ou des substances actives habituellement efficaces sur le champignon pathogène n’arrivent plus à stopper la croissance du champignon pathogène. Le terme « habituellement efficace » est essentiel pour comprendre le concept de résistance aux fongicides. Il implique que quelque chose a changé dans la cible (champignon pathogène) pour réduire l’efficacité du fongicide. Le changement est génétique ce qui implique que le caractère de résistance peut être transmis aux générations futures du champignon résistant. Les souches pathogènes non résistantes sont dites « sensibles » autrement dit « sensibles » aux effets du fongicides. Bien que le terme « insensible » soit parfois utilisé pour décrire la résistance aux fongicides, le terme « résistant » est plus approprié dans le sens où il existe différents degrés de résistance suivant le champignon pathogène et le fongicide. Cet article tente d’introduire le concept de résistance aux fongicides en traitant les principes fondamentaux mais également les incidences pratiques pour l’intendant de pelouses sportives ou d’agrément.
Composantes chimiques et biologiques de la résistance
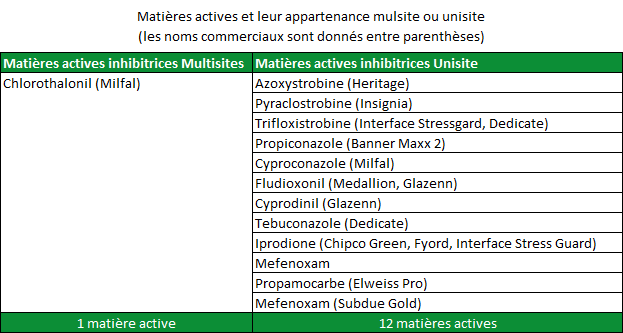
La résistance possède des composantes chimiques et biologiques. La composante chimique est définie par la nature de la substance active inhibitrice. Les fongicides sont classés comme inhibiteurs « multisites » ou « unisites » (tableau 1). Les fongicides multisites figurent parmi les fongicides les plus anciens. Le « Chlorothalonil » est un exemple de substance active multisites très efficace sur un grand nombre de champignons pathogènes du gazon. Une fois pénétré dans la cellule fongique cible, le Chlorothalonil cible des groupes d’atomes constituants de nombreuses protéines essentielles à la physiologie du champignon. Ces protéines régulent des centaines de fonctions métaboliques nécessaires à la croissance de l’organisme. Pour limiter l’efficacité du Chlorothalonil, des milliers de changements simultanés devraient avoir lieu dans les protéines du champignon sans toutefois modifier d’autres processus vitaux. Les chances d’obtenir un tel scénario sont quasiment nulles. En d’autres termes, il est possible de dire que les populations fongiques ne peuvent pas évoluer de manière à devenir résistantes au Chlorothalonil ou tout autre fongicide multisite.
La plupart des fongicides modernes sont inhibiteurs unisites : ils ne perturbent qu’une seule fonction métabolique dans la cible (champignon pathogène). Par exemple, les fongicides dits « DMIs » perturbent la biosynthèse d’un seul composant appelé « ergostérol ». L’ergostérol est un composant essentiel des membranes cellulaires du champignon pathogène. Sans quantité suffisante d’ergostérol, la croissance fongique est stoppée. Des modifications dans la génétique du champignon qui touchent à la fonction inhibitrice de la synthèse de l’ergostérol du fongicide peuvent permettre au champignon de continuer sa croissance, rendant les fongicides de la famille des DMIs moins efficaces voire inefficaces. La probabilité naturelle d’occurrence d’un changement métabolique de ce type est raisonnablement élevée, dans une population fongique, tout particulièrement dans le cas où le champignon se multiplie rapidement et donne lieu à de nombreux individus.
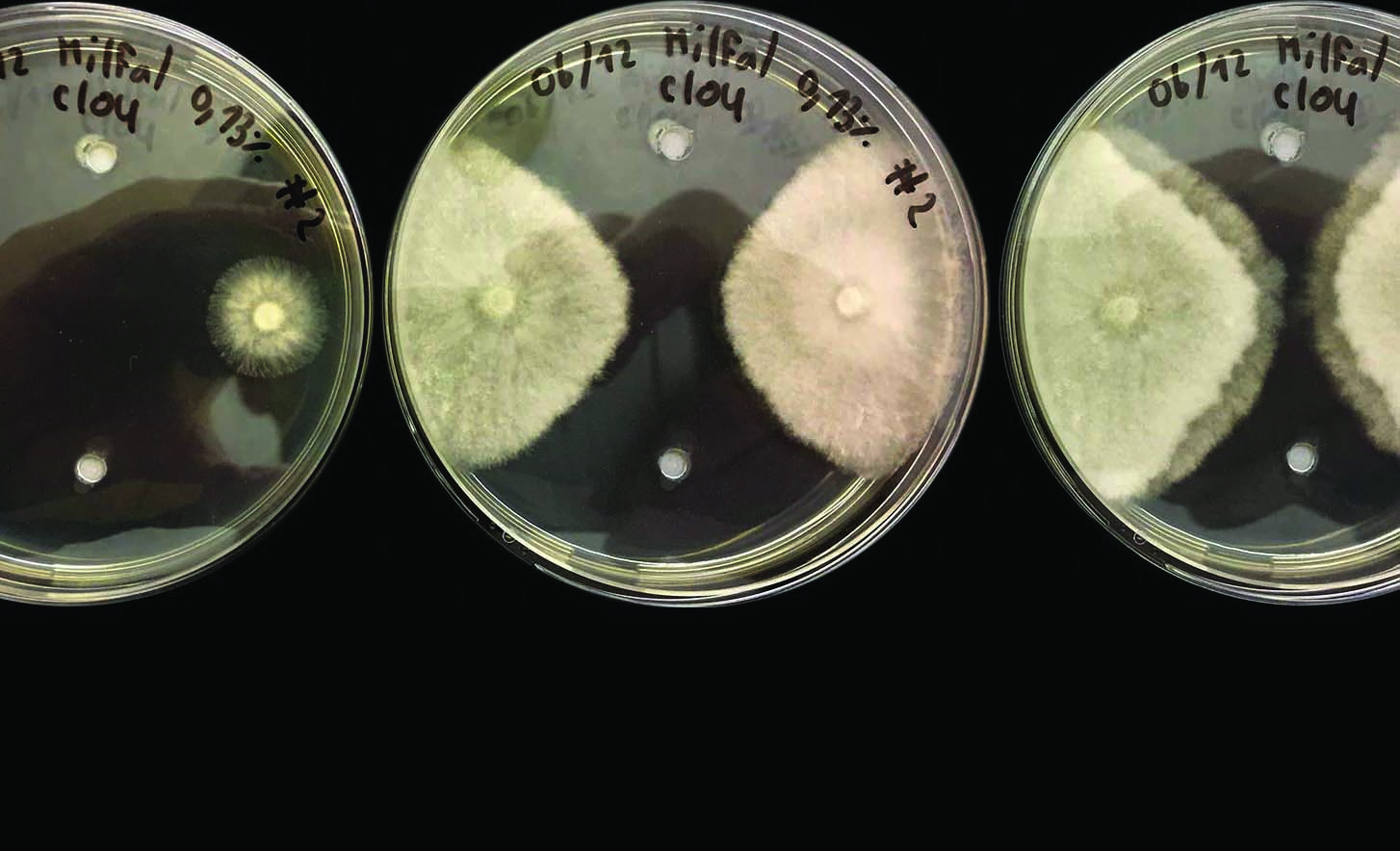
La composante biologique de la résistance est une caractéristique du champignon pathogène. Les problèmes de résistance sont plus fréquents lorsque le champignon est capable de produire de grandes populations. A ce jour, le phénomène de résistance a été identifié sur le terrain pour cinq familles de maladies du gazon : Dollar Spot, Anthracnose, Gray Leaf Spot, fusariose froide et Pythium. Dans la partie Nord-Est des Etats-Unis, le Dollar Spot, causé par le champignon Sclerotinia Homeocarpa est actif durant toute l’intégralité de la saison. Colletotrichum Cereale, pathogène de l’anthracnose sur gazon est également capable de se développer sous de larges gammes de températures. Une seule plante infectée peut produire des centaines de milliers de spores. De la même manière, un grand nombre de spores de Pyricularia Grisea (Gray Leaf Spot) disséminés par le vent sont produits par chaque lésion individuelle. L’agent pathogène responsable de la fusariose froide : Microdochium Nivale est un autre grand producteur de spores qui est actif la partie Nord-Ouest Pacifique des Etats-Unis entre 5 à 9 mois chaque année. Enfin, dans des conditions environnementales idéales, Pythium spp. infecte le gazon avec une rapidité incroyable et un cycle de reproduction très court. Il peut créer une abondance de spores pendant une période de 12 heures seulement. La caractéristique commune à tous ces agents pathogènes réside dans leur capacité à produire rapidement des populations abondantes.
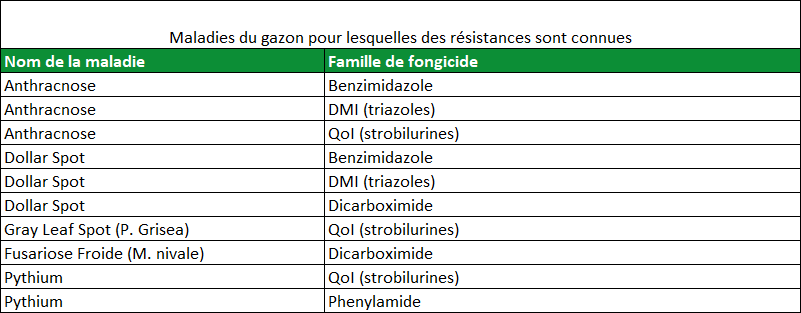
De nombreux champignons pathogènes du gazon sont capables de produire des populations abondantes mais toutes ne sont pas capables de développer une résistance aux fongicides. Le tableau 2 montre les agents pathogènes résistants à certaines familles chimiques de fongicides pour lesquels la résistance est confirmée et étudiée dans de nombreuses publications. Tous les champignons pathogènes listés dans ce tableau produisent des populations massives de cellules infectieuses et toutes les substances actives sont inhibitrices unisites. Alors que de nouvelles substances actives sont introduites sur le marché, nous ne devons pas oublier que tous les fongicides unisites sont vulnérables à l’apparition de résistances au sein des populations fongiques et surtout pour les cinq champignons décrits précédemment. Ce point est très important car les SDHI, nouvelle famille de fongicides unisites, sont homologués sur gazon avec un champ d’action comprenant notamment Dollar Spot et Anthracnose, les deux champignons présentant le risque de résistance le plus élevé.
En fonction de la nature des composantes biologiques et chimiques, l’expression de la résistance peut être appelée « qualitative » ou « quantitative ». L’expression qualitative signifie que la population pathogène est divisée en deux catégories : totalement résistante ou totalement sensible. L’expression quantitative indique que la population pathogène est comprise entre divers degrés de sensibilité au fongicide. Savoir si la résistance est quantitative ou qualitative est important d’un point de vue pratique. Sur le terrain, la résistance quantitative peut être illustrée à travers l’exemple des fongicides de la famille des DMIs (en français IBS : les triazoles) contre Dollar Spot. Elle est caractérisée par une érosion graduelle de la sensibilité aux fongicides de cette famille. Un fongicide ne perd pas complètement sa capacité à contrôler le développement du champignon pathogène et un contrôle satisfaisant de la maladie peut être obtenu avec des doses d’application ou fréquence de traitement plus élevées. La résistance qualitative est quant à elle caractéristique des benzimidazoles (méthyl-thiophanate) avec les populations de Sclerotinia Homeocarpa par exemple. Lorsque les souches pathogènes sont résistantes, l’augmentation des doses ou fréquence d’applications ne donne pas lieu à un meilleur contrôle du champignon.
Résistances croisées et multiples
Résistance croisée
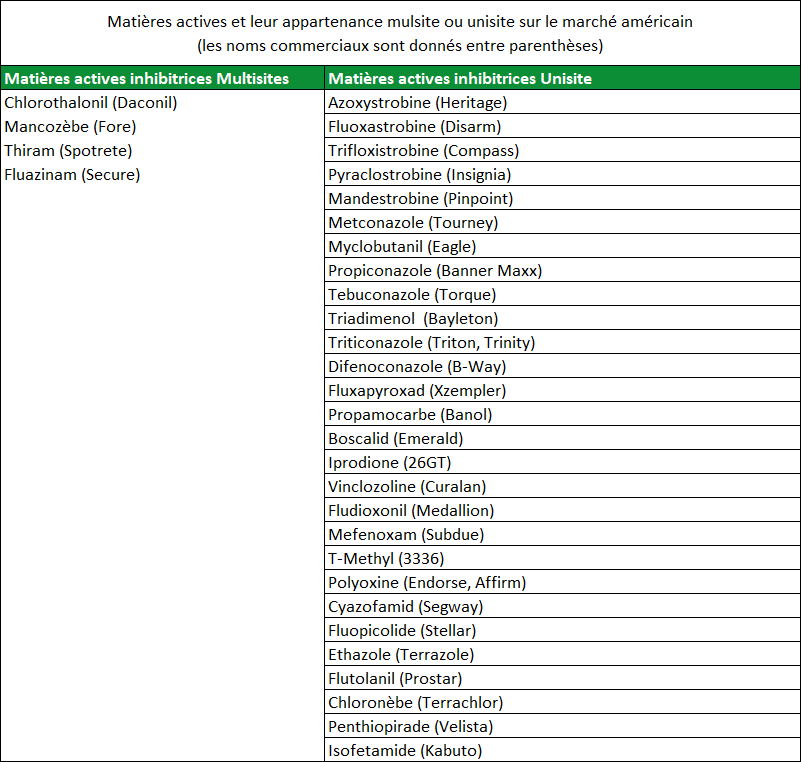
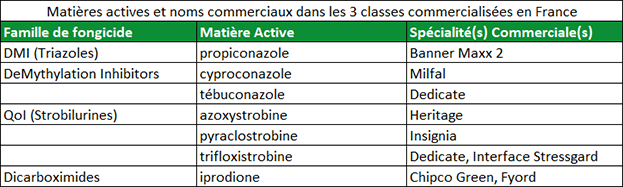
Le terme « mode d’action » est couramment utilisé lorsque l’on parle des fongicides. Le mode d’action correspond à l’interaction entre le fongicide et l’agent pathogène. Les fongicides dotés du même mode d’action perturbent la même fonction métabolique chez la cible pathogène. Par exemple, tous les DMIs (triazoles) possèdent le même mode d’action : ils interviennent tous et de la même manière sur la biosynthèse de l’ergostérol (IBS : Inhibiteur de la Biosynthèse des Stérols). Le mode d’action du fongicide est un paramètre important car quand un champignon pathogène développe une résistance à une matière active dans une famille de fongicides, il y a de fortes chances pour que celui-ci développe également une résistance aux autres matières actives de la même famille. La résistance à une famille de fongicide s’appelle « résistance croisée » (Figure 2). Le tableau 3 répertorie des matières actives avec leur famille d’appartenance. Il est important de souligner à nouveau qu’une population pathogène qui développe une résistance à une matière active d’une famille chimique définie risque fortement de développer une résistance à l’ensemble des matières actives de cette même famille.
Résistance multiple
Lorsque des matières actives unisites provenant de deux ou plusieurs familles de fongicides sont utilisées pour contrôler une maladie, le risque de résistance aux plusieurs familles est très probable. La tendance naturelle d’une population fongique réside dans sa faculté d’adaptation face aux menaces existantes (dans notre cas : les fongicides). Là où des fongicides unisites de différentes familles ont été utilisés contre certains champignons pathogènes, une résistance multiple à plusieurs familles de fongicides a pu se développer. Dans le tableau 4, des cas de résistance multiples à différentes familles de fongicides unisites ont été répertoriés. Le principal problème avec ce type de résistance n’est pas de savoir si elle va arriver mais à quelle vitesse et quelles mesures peuvent être prises pour la ralentir.
Le développement de la résistance est un processus à deux étapes
Une mutation (autrement dit une modification de l’ADN du champignon pathogène qui lui permet de se développer et infecter le gazon en présence de fongicide) est la première étape dans le développement de la résistance aux fongicides. Les scientifiques pensent aujourd’hui que l’apparition d’un individu résistant à un fongicide dans une population fongique est un phénomène naturel mais rare. De ce fait, la probabilité qu’une résistance se développement au sein d’une population dépend de la taille de cette population. Comme expliqué précédemment, les populations fongiques fortes ont ainsi plus de chance de développer une résistance par le biais de cette modification génétique. La loterie est une bonne analogie. En prenant seulement quelques tickets de loterie (représentant une petite population) la chance de gagner un lot est très faible. En prenant plusieurs dizaines ou centaines de tickets (qui représentent alors une forte population) les chances de gagner un lot sont alors bien plus élevées. En rapport aux maladies du gazon, mettre en place une stratégie préventive pour maintenir de faibles populations de champignons pathogènes réduit la probabilité de développement de souches résistantes aux fongicides utilisés. A l’inverse, le risque de développer des souches résistantes est plus élevé si de larges populations fongiques sont présentes. En d’autres termes, ne faire que des applications curatives de fongicides (c’est-à-dire une fois les symptômes visibles) sans aucune application préventive implique de fortes populations pathogènes et un risque élevé de développement de résistance aux fongicides utilisés.
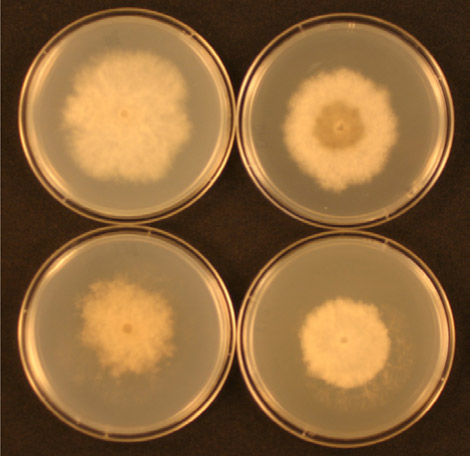
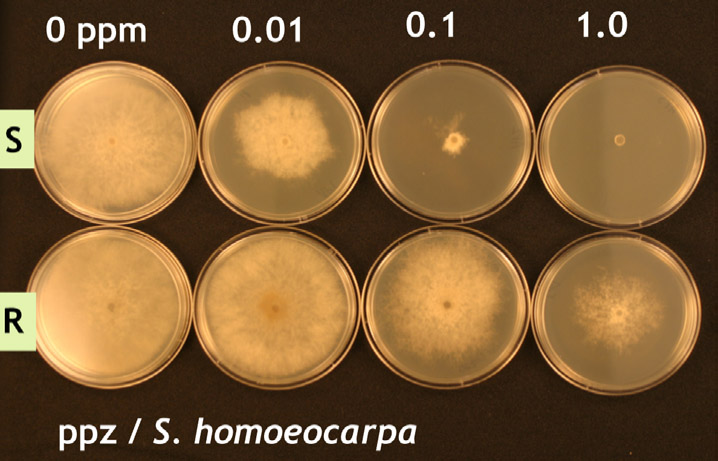
Ainsi, la première étape du processus de résistance est la mutation, un phénomène naturel. La seconde étape provient cependant une action humaine : la pression de sélection. Une cellule fongique résistante n’est pas un problème à elle seule. En réalité, son existence peut passer inaperçue. Le problème a lieu lorsque cet individu se reproduit, permettant à une population pathogène de devenir peu à peu résistante. In fine, l’évolution d’une population résistante à un fongicide est une fonction de la pression de sélection. La pression de sélection augmente lorsque des fongicides de la même famille sont appliqués régulièrement ou systématiquement. Quand un fongicide est appliqué sur une population de champignon pathogène dans laquelle sont présents quelques souches résistantes, les souches sensibles sont éliminées et les souches résistantes « sélectionnées » ou « favorisées » puisqu’elles continuent de se développer (voir figure 3). Par conséquent, des applications de ces fongicides avantagent les souches résistantes, leur permettant alors de devenir dominantes dans la population. L’efficacité du fongicide utilisé ou des fongicides de la même famille diminue alors.
La pression de sélection influence la vitesse à laquelle la population pathogène évolue vers un type d’individus qui sont majoritairement résistants au fongicide où à la famille de fongicides utilisés. Sans aucun doute, la manière la plus rapide de développer une population fongique résistante est d’appliquer exclusivement des fongicides de la même famille. Durant les années 80 et 90, cette erreur a souvent été faite avec l’utilisation systématique des DMIs (triazoles) pour contrôler le développement du dollar spot. Fort heureusement, il est possible d’espérer que les erreurs du passé ne seront plus répétées avec la dernière famille de fongicides unisites : les SDHIs. Syngenta est d’ailleurs sur le point de mettre sur le marché un nouveau fongicide à base de SDHI.
Persistance des populations fongiques résistantes
Le processus à deux étapes de la résistance aux fongicides implique le développement de populations fongiques résistantes aux applications répétées de fongicides unisites de la même famille. Etant donné l’avantage compétitif des souches résistantes sur les souches sensibles en présence d’un fongicide, y’a-t-il à l’inverse une diminution du nombre d’individus résistants en cas d’absence de ce fongicide ? En d’autres termes, en supprimant l’avantage compétitif de la souche résistante (absence d’application du fongicide qui sélectionne cette souche), y’a-t-il un retour des populations vers la situation originale avec des souches majoritairement sensibles ? C’est une question très importante d’un point de vue pratique car dans ce cas, le fongicide auquel la population est devenue résistante est à nouveau efficace plus tard, si celui-ci n’est plus utilisé. La réponse dépend du fongicide, du champignon pathogène et du mécanisme de résistance à l’échelle moléculaire. Etudier la nature fondamentale de la résistance aux fongicides permet un éclairage capital dans la compréhension de la persistance dans le temps des souches résistantes.
Mécanismes moléculaires de la résistance
Pour qu’un fongicide unisite fonctionne, la molécule du fongicide doit « s’imbriquer » parfaitement dans la molécule cible fongique (protéine) comme dans un puzzle en trois dimensions. Lorsque les molécules s’imbriquent, la fonction enzymatique associée au sein du champignon est interrompue : la molécule fongicide prend la place d’une molécule naturelle qui a un rôle vital bien précis dans le métabolisme du champignon. La croissance du champignon est stoppée, le gazon récupère : la maladie est contrôlée. Lorsque qu’une résistance se développe, un changement génétique induit une modification du site d’échange où la molécule fongicide s’imbrique. Cette modification empêche la molécule fongicide de se lier à l’enzyme cible qui peut alors remplir alors son rôle naturel : le champignon pathogène continue son développement. Lorsqu’un changement de ce type a lieu, augmenter la dose ou la fréquence d’application de fongicide n’aura aucun effet puisque la géométrie du site d’échange a été modifiée et le fongicide ne peut plus s’imbriquer. Ce mécanisme de résistance est appelé « mutation du site cible » et il n’y aucune chance que l’absence d’application du fongicide qui pose des problèmes de résistance permette de revenir à des populations initiales sensibles. Parce que ces individus mutants et résistants au fongicide n’ont aucun désavantage compétitif en l’absence de ce fongicide, ils persisteront dans la population au fil du temps.
D’autres mécanismes de résistance existent : la surexpression et l’efflux actif. Dans le cas de la surexpression, les enzymes cibles du pathogène ne sont pas modifiées complètement. Le champignon produit de plus des quantités très importantes d’enzymes cibles de façon à ce que les molécules fongicides ne soient plus présentes en quantité suffisante pour avoir un impact suffisant sur le développement du champignon pathogène. L’efflux actif d’autre part, est une situation dans laquelle certaines cellules fongiques éliminent rapidement les molécules fongicides avant qu’elles s’accumulent à des concentrations létales. Bien que la géométrie des enzymes cibles ne soit pas modifiée, l’efflux actif est un mécanisme au sein du pathogène qui prévient l’accumulation de molécules fongicides à des concentrations empêchant le bon développement du champignon pathogène.
Lorsque la résistance est une fonction de la surexpression ou de l’efflux actif et bien sûr selon les recommandations de chaque produit sur l’étiquette, un contrôle partiel du champignon peut être obtenu en augmentant les doses ou la fréquence d’application (figure 4). Les études en laboratoire sur les mécanismes de résistance appuyé par un monitoring de la persistance des populations sensibles ou résistantes sur le terrain sont rares. Cependant, plusieurs études menées sur les fongicides et champignons pathogènes des grandes cultures suggèrent que dans les cas autres que la « mutation du cite cible » (voir plus haut), l’absence d’application de fongicides ayant donné lieu à une résistance sur un certain laps de temps, mène à un retour vers des populations initiales sensibles à ce même fongicide. Bien que la plupart de ces études aient été menées sur les champignons pathogènes en grande culture, certaines conclusions peuvent être extrapolées au gazon puisque de nombreuses matières actives sont utilisées dans les deux domaines.
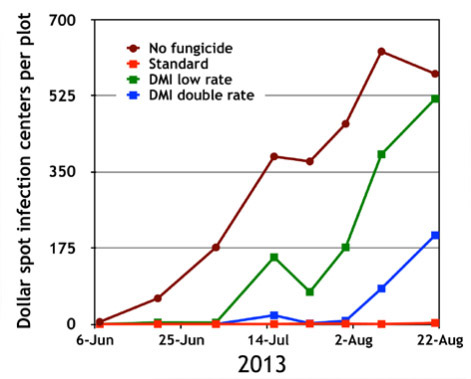
Etude de cas : fongicides pour contrôler les maladies du gazon
Benzimidazoles (méthyl-thiophanate)
Des preuves empiriques et expérimentales montrent que les phénomènes de résistance aux benzimidazoles pour les champignons Sclerotinia Homeocarpa et Colletotrichum Cereale sont le résultat d’une mutation du site cible. Ces mêmes résultats vont dans le sens d’un non-retour aux populations sensibles initiales en cas d’absence de fongicide de cette famille. Ces hypothèses sont appuyées par de nombreuses études sur pathogènes des grandes cultures qui indiquent que les populations résistantes persistent dans le temps.
Fongicides QoI (strobilurines)
Des mutations du site cible ont mené à des phénomènes de résistance aux strobilurines pour le champignon pathogène « Gray Leaf Spot » (Pyricularia Grisea). Des études scientifiques ont montré que la résistance se maintient sur de nombreuses générations de champignons pathogènes résistants, confirmant l’absence de retour aux populations sensibles initiales en absence de fongicides de cette famille. Des phénomènes de résistance croisée ont également été observés. La résistance d’une souche à une strobilurine implique donc une possible résistance à l’ensemble des molécules de cette famille. La résistance aux strobilurines pour l’anthracnose (Colletotrichum Cereale) est dûe à une mutation du site cible, comme le montrent différentes études menées sur des pathogènes en grandes cultures. Bien que des variations existent au sein des populations résistantes à cette famille de fongicides, des évidences certaines vont également dans le sens d’un non-retour aux populations initiales sensibles en l’absence d’application. Les populations résistantes persistent donc dans le temps.
Dicarboximides (iprodione)
Les souches de Dollar Spot résistantes à l’iprodione ont généralement une croissance plus faible que les populations sensibles selon de nombreuses observations. Il est donc fortement probable que cette résistance donne lieu à un retour vers des populations sensibles initiales (les individus sensibles ont une croissance plus forte que les individus résistants). Des résultats similaires ont été observés dans le cas de champignons pathogènes résistants aux dicarboximides. Pourtant, il existe peu d’études sur le terrain menées sur les pathogènes du gazon. Dans une étude menée au champ sur le Botrytis (pathogène virulent de la vigne en Europe), l’utilisation de dicarboximides a été stopée suite au développement de résistance du champignon à cette famille. Après quelques années, de nouvelles applications de ces produits ont à nouveau été réalisées dans un plan de protection alternant différentes familles (une application seulement de dicarboximide par saison) avec succès. Les mécanismes de développement de résistance aux fongicides de cette famille ne sont pas encore très clairs. Certaines études suggèrent une mutation du site cible alors que d’autres suggèrent plutôt l’efflux actif comme origine de la perte d’efficacité.
Fongicides DMIs (triazoles)
Trois mécanismes moléculaires à l’origine des résistances aux DMIs ont été identifiés chez plusieurs champignons pathogènes. Des recherches récentes sur Dollar Spot suggèrent que les phénomènes de mutation du site cible et efflux actif sont impliqués. L’efflux actif expliquerait pourquoi des doses élevés appliquées sur des intervalles courts contrôlent les champignons pathogènes, surtout lorsque la pression du parasite est faible. Des études au champ menées sur des populations de Dollar Spot résistantes aux DMIs montrent tout de même que la proportion d’individus résistants reste élevée même si un retour à des populations sensibles est observé en l’absence d’application. Sachant que des cas de résistance croisée ont été observés et que deux régulateurs de croissance aux effets fongistatiques similaires aux DMIs sont régulièrement utilisés par les greenkeepers (flurprimidol et paclobutrazole), il est clair que l’efficacité des DMIs est très limitée dans le cas d’apparition de souches résistantes.
Fongicides SDHI
Le phénomène de résistance aux SDHI a été étudié pour de nombreux champignons pathogènes des grandes cultures. Dans ces cas, les mécanismes de résistance associés concernent de multiples mutations des sites cibles. Les populations résistantes se développent rapidement dans le cas d’utilisation de SDHIs comme le boscalid, le fluopyram, le fluxapyroxad et le penthiopyrad. Des cas de résistance croisée existent et sont influencés par le type de mutation et le type de matière active. Dans tous les cas de résistance, le retour aux populations sensibles initiales n’a pas été observé, montrant que les souches résistantes seront présentes sur les prochaines générations. A ce jour, aucun cas de résistance aux SDHIs n’a été signalé pour le gazon. Pourtant, le développement de résistance est fortement probable si la pression de sélection augmente par le biais d’une utilisation exclusive de cette famille de fongicide.
Résistance Multiple
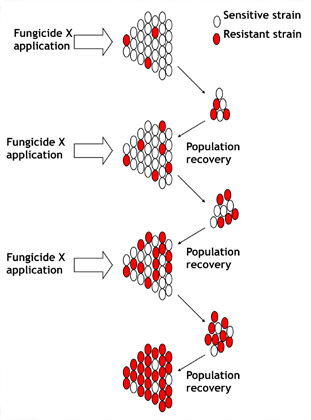
la survie au long terme de souches de Dollar Spot ou Anthracnose multi-résistantes est influencée par les mécanismes impliqués et l’utilisation continue de fongicides devenus inefficaces. La capacité de survie de souches multi-résistantes reste incertaine. Les études menées sur pathogènes des grandes cultures qui suivent précisément la fréquence d’apparition de souches résistantes et sensibles sur de nombreuses années estiment, dans une certaine mesure, le retour au fil du temps vers des populations sensibles en l’absence d’application des fongicides posant problème. En d’autres termes, les populations de champignons pathogènes résistants diminuent avec le temps en l’absence de fongicides à risques mais ne disparaissent jamais complètement de la population. Ce phénomène de retour vers des conditions initiales permet l’utilisation ultérieure de fongicides devenus inefficaces lorsque ceux-ci sont utilisés dans une rotation réfléchie des familles et des matières actives. Ces fongicides ne doivent alors plus être utilisés pendant au moins un an et seront utilisés par la suite au maximum une fois par saison.
Une approche pratique de la résistance aux fongicides
Une compréhension des mécanismes de résistance aux fongicides pourra éventuellement guider les utilisateurs pour le contrôle des champignons pathogènes du gazon. Jusque-là, le contrôle des maladies devrait être basé sur une stratégie simple pour gérer le processus en deux étapes à l’origine du phénomène de résistance. Du fait d’une plus forte probabilité de résistance dans le cas de populations dynamiques de pathogènes, les stratégies permettant de maintenir les populations peuvent aider à prévenir tout risque de résistance. Les solutions alternatives sont également une solution efficace pour maintenir de faibles populations. Les traitements préventifs sont donc à encourager et spécialement les programmes basés sur des fongicides multisites. Les fongicides contenants des matières actives multisites sont tout particulièrement intéressants puisqu’ils diminuent la probabilité d’apparition de populations pathogènes résistantes. Le Chlorothalonil et le Fluazinam sont des exemples de matières actives multisites pour le contrôle du Dollar Spot. Limiter l’utilisation de matières actives unisites ou mélanger ces matières actives avec des matières actives multisites permet de réduire efficacement le risque d’apparition de résistance. Finalement et c’est la conclusion la plus importante : il est impératif d’éviter d’utiliser exclusivement une seule famille de fongicides pour contrôler une maladie. Avant que des conclusions expérimentales prouvent le contraire, alterner les différentes familles de fongicides est la solution la plus efficace.
Bibliographie
- Amiri, Achour, et al. (2014) “Resistance to fluopyram, fluxapyroxad, and penthiopyrad in Botrytis cinerea from strawberry.” Plant Disease, vol. 98, no. 4, pp. 532-539.
- Hahn, Matthias. (2014) “The rising threat of fungicide resistance in plant pathogenic fungi: Botrytis as a case study.” Journal of Chemical Biology, vol. 7, no.4, pp. 133-141.
- Hulvey, Jon, et al. (2012) “Overexpression of ShCYP51B and ShatrD in Sclerotinia homoeocarpa isolates exhibiting practical field resistance to a demethylation inhibitor fungicide.” Applied and Environmental Microbiology, vol. 78, no. 18, pp. 6674-6682.
- Kim, Y. S., et al. (2003) “Field resistance to strobilurin (QoI) fungicides in Pyricularia grisea by mutation in the mitochondrial cytochrome b gene.” Phytopathology, vol. 93, no. 7, 2003,pp. 891-900.
- Latin, Richard. (2011) A Practical Guide to Turfgrass Fungicides. APS Press
- Ma, Z., and Themis Michailides. (2005) “Advances in understanding molecular mechanisms of fungicide resistance and molecular detection of resistant genotypes in phytopathogenic fungi.” Crop Protection, vol. 24, no. 10, pp. 853-863.
Traduction : R. GIRAUD
Cet article est téléchargeable au format *.pdf en cliquant sur le lien suivant.
Copyright United States Golf Association©. Tous droits réservés



{kind=link}